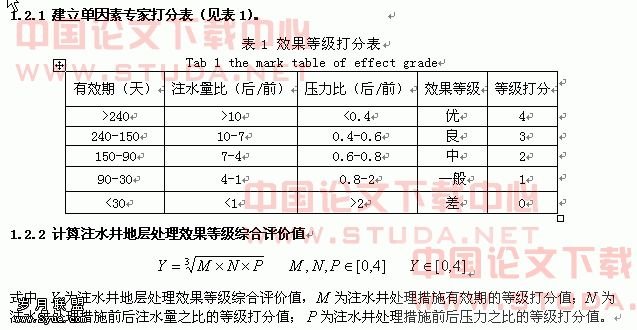
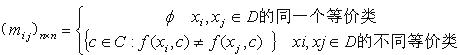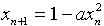植物细胞凋亡
来源:岁月联盟
时间:2010-07-11
论文摘要: 细胞凋亡又叫细胞程序性死亡,是植物正常发育中必不可少的一部分,目前已成为植物细胞生物学研究的一个热点。本文对植物凋亡的一般特征、植物营养和生殖生长中的细胞凋亡以及植物-病原物互作中的细胞凋亡进行了综合评述,并对植物细胞凋亡研究的现实意义进行了探讨。
细胞凋亡是多细胞生物体在生理或病理条件下部分细胞所采取的一种由内在基因编程调节,通过主动的生化过程而自杀死亡的方式[1 ]。由于细胞凋亡受到严格的由遗传机制决定的程序性调控,所以常常又称为细胞编程性死亡。细胞凋亡的现象最早是Kree在1965年观察到的,经过进一步深入研究之后,他于1972年将其重新命名为细胞凋亡。之后近20年,细胞凋亡的研究主要集中在动物,人们越来越认识到细胞调亡在动物生长发育中、尤其在维持动物体内细胞和组织平衡、特化、形态建成和防病、抗病过程中的重要作用。同动物一样,在植物生长发育中也存在着细胞凋亡现象。但由于植物生长发育和细胞结构的特殊性,有关植物细胞凋亡的研究起步较晚。近年来,随着植物细胞凋亡的研究进展,人们逐渐认识到细胞凋亡是高等植物生长发育的必要组成部分,同时也是植物体度过不良环境的重要手段。目前,植物细胞凋亡的研究已成为近年来植物细胞生物学的新兴研究领域和热点之一。本文就植物细胞凋亡的一般特征、检测方法、在植物中的存在及意义作一综合阐述。
1 植物细胞凋亡的一般特征
经历细胞凋亡过程的细胞呈现一些典型的形态学变化,光学显微镜或显微镜观察可见:细胞体积缩小,染色质凝集、断裂、趋边化,细胞器解体、消失,细胞膜发泡形成凋亡小体(其中包含有凝集的细胞核断片和细胞器) [3.4]。随着研究的深入,分子生物学证据也逐步被阐明:细胞染色质DNA 在核小体连接部位断裂,其片段大小为200bp的倍数,经琼脂糖凝胶电泳可见到特征性的DNA 梯度(DNA ladder) ,此特征还可以通过超速离心、末端标记电泳以及原位缺口翻译技术等进行定性、定量测定。细胞形态学和分子生物学的变化是细胞凋亡的重要诊断依据。
2 细胞凋亡的检测方法
2. 1 细胞形态学观察法
苏木素- 伊红(HE) 染色法: 石蜡切片的HE 染色是组织形态学检测的常规方法, 光学显微镜下细胞核呈蓝黑色, 胞浆呈淡红色。凋亡细胞在组织中单个散在分布, 表现在核染色质致密浓缩, 核碎裂等。
(1)电子显微镜。电镜观察,凋亡细胞染色质固缩,常聚集于核膜上呈境界分明的块状或新月形小体, 初期细胞可见完整的细胞器, 细胞膜完整, 凋亡小体形成。目前一致认为, 电镜下获得凋亡细胞特征性的形态学改变是判断细胞凋亡的最可靠依据。
(2) 荧光显微镜。对体外培养的活细胞经荧光色素处理, 可在荧光显微镜下观察细胞形态改变。常用荧光色素有吖啶橙、Hoech st 33258或Hoech st 33342、碘化丙啶(P I)、溴乙锭(EB)。前两种可分别进入活细胞和死细胞, 而后两种荧光素仅能进入死细胞。不同的荧光素使核着染不同颜色的荧光, 正常细胞呈均匀荧光染色, 而凋亡细胞呈致密浓染的颗粒状或块状荧光。
2. 2 反映凋亡细胞膜改变的方法:染料排斥法。
除了电镜能反映细胞膜完整性外, 还可用染料排斥法, 如台盼蓝、P I 等。坏死细胞膜破损, 被染料着染。而凋亡细胞细胞膜完整, 不被着染。但在体外培养的细胞最终也会发生继发性坏死。因此, 此法不能单独用来判断凋亡细胞。另一种方法是判断胞质膜的不对称性。在正常细胞膜上, 磷脂酰丝氨酸基团(PS) 位于胞内侧, 而在细胞凋亡早期膜上此基团则转向胞外侧, 以利于被吞噬。因此, 磷脂酰丝氨酸基团位置的改变, 可作为凋亡细胞的一个标志。
2. 3 反映脱氧核糖核酸有断裂的方法
细胞凋亡过程中,DNA有规律地断裂可以通过下述几种方法检测出来。
(1) 琼脂糖凝胶电泳法。细胞悬液经裂解消化按常规法提取DNA后, 于含EB 的琼脂糖凝胶中进行电泳,正常细胞DNA呈单一条带。细胞凋亡时呈典型的梯状条带,系180~ 200 bp 左右的及多聚核小体的梯状DNA条带。坏死时则呈现模糊的弥散状条带。DNA电泳法是判断细胞凋亡的经典方法. PEG6000 诱导的小麦叶片[7] 、羟自由基诱导的烟草细胞[8] 、细胞色素c诱导的胡萝卜和烟草原生质体[9] 和乙烯诱导的胡萝卜原生质体[10]发生PCD 时均检测到DNA 梯状条带。
(2) 流式细胞仪检测法。细胞发生凋亡时, 其细胞膜的通透性增加, 但其程度介于正常细胞和坏死细胞之间, 利用这一特点, 被检测细胞悬液用萤光素染色利用流式细胞仪测量细胞悬液中细胞萤光强度来区分正常细胞、坏死细胞和凋亡细胞。
( 3 ) 原位末端标记法( In Situ End2L abeling,ISEL )。通过DNA 多聚酶I 把已标记的核苷酸结合到DNA 的单链断裂处, 以寻找有无Ap 发生。标记的方法有同位素标记、荧光素标记、地高辛或生物素标记等。
(4) 原位切口平移法( In Situ N ick T ran slat ion, IS2N T )。利用DNA 多聚酶将核苷酸整合到Ap 细胞内断裂的DNA 3′羟基末端, 同时水解5′末端,以修复DNA。若用已标记的核苷酸, 即可显示出有断裂DNA 的细胞。该法同样也可用于细胞悬液中Ap 的观察。
( 5) 末端转移酶介导的缺口末端标记法(TdT 2m e2diated X2dU TP n ick end labeling, TUN EL )。末端转移酶(TdT ) 介导的X2dU TP 缺口标记法是目标原位检测Ap 最为敏感、快速、特异的方法, 其具有广泛的应用前景。末端转移酶(TdT ) 可催化在DNA 片段的3′羟基末端合成多核苷酸聚合物的反应, 即DNA 片段加尾。利用末端转移酶(TdT ) 将标记的脱氧核苷酸转移到DNA 缺口或3′羟基末端上, 通常所用的核苷酸为dU TP, 标记物为地戈辛、生物素、荧光素等。
(6) EL ISA 法。对Ap 细胞内DNA 片段的检测还可用EL ISA 法。悬浮细胞经裂解, 高速离心去除核的成分后, 取上清加入已包被有抗组蛋白抗体的反应板, 反应后再加酶标抗DNA 抗体, 若上清中含断裂的DNA片段, 则可通过此双抗体夹心法得以检出[11]。
3 植物发育过程中的细胞凋亡
萌发的种子中的糊粉层、维管束的木质部、生殖器官的组织(如花药和子房)及根冠等组织中均有细胞凋亡的发生[12]。虽然在细胞水平上, 与细胞凋亡相关联的水解酶的激活、一些蛋白的失活以及核DNA的断裂都可以经常观察到, 但是这些现象的发生机制到近来才有所了解。
3.1 导管的形成
导管是由排列有序的死亡的导管分子(tracheary elements, TEs)构成。王雅清和崔克明[ 13 ]对杜仲木质部导管分化的研究证明,其分化过程也发生了细胞凋亡。所有这些研究都表明木质部导管分化与细胞凋亡有密切关系。玉米生长过程中在一定条件下根部皮层细胞崩溃死亡形成通气组织, 而通气组织与植物的同化、呼吸、蒸腾作用都有密切关系[14 ].
3.2 单性花的形成
许多单性花植物在花原基分化时存在雌蕊和雄蕊原基细胞,在后期发育的特定阶段雌蕊或雄蕊原基细胞出现细胞凋亡,从而最终形成单性花。
3.3 大、小孢子的形成和发育
大多数种子植物中, 大孢子母细胞减数分裂形成4 个大孢子。仅有1 个能发育成雌配子体, 其余的3个大孢子退化。例如, 蕨类植物大孢子母细胞减数分裂产生4个大孢子, 这4个大孢子通常呈线型或T型排列, 仅有1个能继续发育成雌配子体, 其余3个都死亡。对其超微结构的研究表明,其退化解体过程也符合细胞凋亡的基本特征[15] 。
3.3 雌雄配子体的发育
植物中雌雄配子体的发育有细胞凋亡参与其中。裸子植物雄配子体发育过程中, 原叶细胞的退化和雌配子中颈细胞、腹沟细胞的消失及珠心细胞的衰退也是细胞凋亡的结果。在被子植物雌配子体(胚囊)发育过程中,珠心组织被作为营养物质吸收而退化的过程是细胞凋亡[16].
3.4 胚的发育
在胚性细胞分化和发育过程中,存在着细胞凋亡[17]。植物的胚由受精卵发育而成, 在胚的形成过程中,助细胞、反足细胞和胚柄细胞都因发生细胞凋亡而消失。胚柄由受精卵第一次分裂形成,当胚发育到一定阶段,胚柄发生细胞凋亡, 形态上表现为质壁分离, 原生质体固缩。单子叶植物的种子中, 在胚和胚乳之间有一层或几层排列整齐的糊粉层细胞, 含大量糊粉粒。胚胎发育早期由胚柄提供营养形成种子, 后期则通过糊粉层细胞形成分泌组织, 分泌水解酶, 水解胚乳成分, 种子萌发后, 糊粉层功能完成, 便开始凋亡,是典型的细胞凋亡。在种子萌发过程中,其他胚乳和无胚乳种子子叶中一些贮藏细胞也会发生类似的细胞凋亡, 没有这些细胞凋亡,幼苗就不能正常生长发育, 会因饥饿而死亡。
3.5 根冠细胞的死亡
根冠位于根尖的顶部,是由许多薄壁细胞组成的冠状结构。在根的发育过程中,根冠细胞不断脱落,并由顶端分生组织不断产生新的细胞,从内侧补充使根冠细胞得以保持定数。对根冠细胞脱落的研究证明,其脱落过程是典型的细胞凋亡。正是这些细胞的主动死亡,才保证了根顶端分生组织在生长过程中避免与土壤磨擦而受伤,进而保证了根的正常发育。对玉米根尖进行低温胁迫或用细胞毒素类药物如放线菌D、秋水仙碱处理后,这些根尖分生组织细胞同样具有DNA ladder、染色质和细胞核浓缩等特征,说明环境因子和药物也可诱导根尖细胞发生凋亡[19.20] 。
3. 6 叶发育过程中的细胞凋亡
在叶子的发育过程中,叶缘的各种裂、齿和叶片中的空洞(如龟背竹叶片) 的形成等都是由于相关部位细胞的凋亡所造成的。此外,对叶片衰老过程的研究发现,衰老起始时,叶绿体首先被自体吞噬,此后水解酶、RNA 酶等活性上升,而且以液泡内半胱氨酸蛋白酶活性最为显著[21 ] ,这些都是细胞凋亡的特征。因此,叶子脱落前叶片的衰老过程也是PCD。
4 环境胁迫诱导的植物PCD
4. 1 植物超敏反应中的PCD
超敏反应(hypersensitive response HR) 是植物被病原物侵染后所引起的适应性反应,其中的细胞死亡被证明是细胞凋亡。在植物超敏反应中,DNA 片段化,特征性切割核小体的核酸酶被激活等生理生化特征和凋亡小体等形态特征都被证实。 4. 2 盐胁迫诱导的PCD
无机盐KCN、NaCl 、CaCl2 和一些重金属离子等在一定条件下均可诱导植物细胞出现与动物细胞凋亡类似的特征。宁顺斌[22]等人的实验证明烟草、玉米的根尖在高盐(NaCl 500mmol/ L)处理后,出现明显的DNA 梯状电泳图谱。林久生和王根轩[23]用20%PEG溶液(-0.63 MPa)对小麦根系进行渗透胁迫,在小麦叶片DNA琼脂糖凝胶电泳图谱上观察到明显的梯状DNA 条带,表明PEG处理诱发了DNA核小体间的断裂,末端脱氧核糖核酸转移酶介导的3’OH末端标记法(TUNEL)检测出现阳性结果。
4. 3 活性氧与植物细胞凋亡
活性氧是一类具有强氧化能力的物质,主要包括超氧化物、过氧化氢、羟自由基等,各种逆境条件,包括冷害、渗透胁迫、低氧、臭氧、紫外线等导致的植物细胞凋亡最终都与活性氧的产生有关。当细胞外一些信息如辐射、高温等通过细胞活性氧传入细胞引起其脂质过氧化或与细胞凋亡有关基因的表达时,细胞也会凋亡[24]。陈明等[25]研究发现以一氧化氮(NO)供体硝普钠(SNP)处理小麦可以明显提高ROS 清除酶,如过氧化物酶、超氧化物歧化酶、抗坏血酸氧化酶等活性,从而清除因盐胁迫产生的氧自由基或活性氧ROS,或直接清除ROS来保持细胞处于还原状态。
5 研究植物细胞凋亡的意义及展望
导管细胞的退化死亡、筛管细胞原生质的自溶,形成了植物体的输导组织;这些细胞死亡之前,细胞内物质可被其他细胞回收利用,这是植物能够独立营养的一个特性,叶片衰老死亡即是适应营养重新分配的结果,但这个过程却影响了农产品的产量。因此,要搞清植物细胞凋亡的发生程序,对粮食生产及作物储藏技术改良都具有重要的现实意义。在超敏反应中,被病原体感染的宿主细胞采取主动死亡的方式,从而限制感染部位病原菌的生长,阻止病原菌的传播,以达到防病抗病的目的。这种植物自身的主动抗病反应,若在植物抗病育种中加以应用,使植物能够自动、有效地抵抗病原物的侵染,就可以减少农药的使用,避免环境污染,从而提高人类生活质量。
由于植物细胞凋亡的同步性很低、凋亡时间很短,同时由于细胞内各种因子相互作用,调控机制及其复杂,使分子生物学技术应用于细胞水平的研究存在很大困难。近年来,利用非细胞体系来研究细胞凋亡的模式的建立和应用弥补了上述不足。有研究表明[26],利用非细胞体系研究细胞内复杂的生化活动具有独特的优越性,在细胞周期调控、DNA 复制、核小体与染色质构建等研究中发挥了重要作用。非细胞凋亡体系的建立与利用,在很大程度上促进了人们对植物细胞凋亡生化和分子机制的研究,为植物细胞凋亡研究开辟了新途径。
随着植物细胞凋亡的研究的逐步深入,发现植物细胞凋亡需要研究的方面还很多。植物体发生细胞凋亡的机理还不清楚,植物细胞中与细胞凋亡有关的基因研究还远没有动物深入。尽管许多实验表明植物细胞凋亡与动物是相似的,但分子水平共同特征少,目前仅发现少数几个基因参与植物细胞凋亡的过程[27] 。研究过程中常局限于某一特定现象,很少有将这些现象和植物发育的具体过程联系起来,加上植物生长周期较长,给研究带来一定困难。植物细胞凋亡的研究如果能与植物的利用联系起来,将具有重要实践价值。如能发现诱导果实发育中细胞凋亡发生的因子,通过人为调控,改变生长发育期,提高果品产量和品质,则将会极大地推动果树化生产的。
[ 1 ] 张亚历等. 编程性细胞死亡及研究方法[J ] . 细胞生物学杂志,17 (3) :127 - 128.
[2] 蒋争凡,翟中和. 细胞凋亡[J ] . 通报,1999 ,44 (18) :1 920 - 1 928.
[ 3 ] 宝福凯. 编程性细胞死亡的研究现状及展望[J ] . 1995 年,细胞生物学杂志,17 (3) :122 - 126.
[4]杨帆,王文亮,王伯云. 细胞与分子免疫学杂志,1997,13(1):8—9
[7]林久生,王根轩. 渗透胁迫诱导的小麦叶片细胞程序性死亡. 植物生报,2001 ,27 :221~225.
[8]夏慧莉,陈浩4 华志明,陈睦传,沈明山. 生物工程进展,1998,18(3):32—36
[9]孙英丽,赵 允,刘春香,翟中和. 细胞色素c 能诱导植物细胞编程性死亡. 植物学报,1999 ,41 :379~383.
[10] 周 军,朱海珍,姜晓芳,戴尧仁. 乙烯诱导胡萝卜原生质体凋亡. 植物学报,1999 ,41 :747~750.
[ 11 ] 张伟成,严文梅,娄成后. 小麦衰退珠心中解体原生质体向胚囊的迁移及其对增殖中反足细胞的哺育[J ] . 植物学报,1984 ,26 (1) :11221.
[ 12 ] 郭小丁. 植物细胞的编程性死亡[J ] . 1998 年,植物学通报,15 (5) :40 - 43.
[ 13 ] 王雅清,崔克明. 杜仲次生木质部导管分子分化中的程序性死亡[J ] . 植物学报,1998 , 40 : 1 102 - 1 107.
[ 14 ] 华志明,陈睦传,沈明山. 细胞生存与死亡的社会控制[J ] . 1998 年,生物工程进展,18 (3) :32 – 36
. [15 ] 陈朱希昭,陈耀堂,高信曾. 太谷核不育小麦花药组织和小孢子发生的超微结构研究[J ]1 植物学报,1984 , 26 :235 – 240
[ 16 ] 尤瑞麟. 小麦珠心细胞衰退过程的超微结构研究[J ] . 植物学报,1985 , 27 : 345 - 353.
[ 17 ] 邢更妹,李杉. 植物体细胞胚发生中抗氧化系统代谢动态和细胞程序性死亡[J ] . 生命科学,2000 ,12 (5) :214 - 216..
[ 19 ] 宁顺斌,宋运淳,王玲,等. 低温胁迫诱导玉米根尖细胞凋亡的形态和生化证据[J ] . 植物生理学报,2000 ,26 (3) :189 - 194.
[ 20 ] 宁顺斌,宋运淳. 药物诱导的玉米根尖细胞凋亡[J ] . 植物学报,2000 ,42 (7) :693 - 696.
[ 21 ] 杨征,蔡陈凌,宋运淳. 植物细胞凋亡研究进展[J ] .生物化学和生物物理进展,1999 ,26 (5) :4392443
[22] 宁顺斌,宋运淳,王玲等. 盐胁迫诱导的植物细胞凋亡———植物抗盐的可能性生理机制[J ] . 实验生物学报,2000 ,33 (3) :6132623.
潘建伟, 陈虹, 顾青 (2002) 环境胁迫诱导的植物细胞程序性死亡. 遗传, 2 4 : 385-388
[23] 林久生, 王根轩. 渗透胁迫诱导的小麦时片细胞程序性死亡. 植物生理学报 , 2001, 27(3): 221~225
[24]吴 逸,戴尧仁. 羟自由基诱导烟草细胞凋亡.植物生理学报,1999 ,25 :339~342.
[25] 陈 明, 沈文飚, 阮海华, 等. 一氧化氮对盐胁迫下小麦幼苗根生长和氧化损伤的影响. 植物生理与分子生物学学报, 2004, 30(5): 569~576
[26] 孙英丽,赵允,翟中和. Cytochrome C 能诱导植物编程性死亡[J ] . 植物学报,1999 ,41 :379 - 387.
[27]Hua ZM(华志明) , Chen MC(陈睦传) , Shen MS (沈明山) .1998. Programmed cell death during plant growth and devel2opment (植物生长发育中程序性细胞死亡) [J ] . A dvanceson Bioengineering (生物工程进展) , 18 (3) : 32 - 36.
下一篇:微生物肥料的作用效果及广阔前景