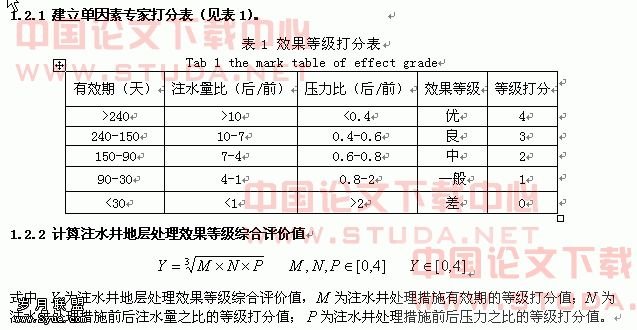
担子菌和子囊菌交配型基因的研究进展
来源:岁月联盟
时间:2010-07-11
关键词:交配型基因 子囊菌 担子菌 酵母菌 性亲和性 在真菌的有性生殖过程中,雄性孢子的交配型选择是至观重要的,不论是在丝状真菌还是在非丝状真菌中,交配型都能说明真菌的遗传控制情况和有性生殖的过程。也就是说,在整个真菌有性生殖过程中交配性基因对真菌的性别控制和遗传进化起着决定性作用。现在的研究大都以子囊菌和担子菌为对象,所以这两个亚门的某些菌种大都研究比较清楚。对酵母菌交配型基因的研究(主要以Sannharomyces cerebvisiae 和Sannharomyces pombe为材料)有许多的新发现。研究还发现,酵母和丝状真菌中的交配决定因子有着十分相似的控制机制[1],但是功能却不相同。研究担子菌和子囊菌的代表菌种,主要从交配反应、交配所需的细胞组分、以及交配型基因的结构和功能等方面着手。由于技术方面的限制,使得目前对接和菌纲和壶菌纲中物种的交配型研究较少。交配型基因是性亲和表现型遗传决定因子。这类真菌主要是是通过异宗配合进行有性生殖的,它们自交不育,必须由两个从不同的单核孢子发育而来的、性亲和的菌丝体配合才能形成双核体,进而发育出子实体。真菌群体中交配型因子的研究,不仅可以加深相关遗传背景的了解,而且可以为菌种鉴定提供一个良好的标准。本文从子囊菌和担子菌具体个例研究的情况来对近几年来对真菌中的交配性基因的研究做一综述。
1交配型基因的结构与命名1.1担子菌种交配型基因结构与命名担子菌通常有性生殖通过不同交配型的营养菌丝间或一个粉孢子同营养菌丝的一个细胞间的融合进行,发生典型的体细胞配合(somatogamy)。大多数担子菌是异宗配合进行有性生殖的,该过程有一个或两个遗传因子决定,由一个因子决定交配型的机制称为二极性交配系统;由两个独立分配的因子控制的交配称为四极性交配系统。二极性交配系统的遗传结构相对简单,A因子是唯一的控制因子,它实际上由两个紧密连锁的基因簇组成,四极性交配系统中, A和 B因子分别对应位于不同染色体上的两个交配型位点(Mating type locus),两个单核菌丝相遇,只有当它们形成AαAβBαBβ型时,才能产生具有锁状联合的双核体,完成整个有性生活史。A位点的交配型基因编码两类含相似同源结构域的蛋白质,两种蛋白质形成异源二聚体,调节依赖A因子的发育过程。 A交配位点有两个紧密连锁的基因复合物Aα和Aβ组成,包括四对基因,分别成为a、b、c和d基因对,a和b之间有7kb的序列将它们分为α和β两个复合物,与遗传学上定位的Aα和Aβ是对应的[2],α复合物有一个基因对a ,β复合物含有b、c和d三个基因对。每个位点有多个复合等位基因,Aα有9个等位基因,Aβ有32个等位基因[3]。B交配位点的交配型基因编码信息素及其受体,主要控制核的迁移。担子菌的B交配型位点与A位点一样,都是由a和β两个基因复合物(基因簇)组成,目前在裂褶菌中已分离到了Bα1和Bβ1复合物,它们的结构很相似,都编码1个信息素受体基因( Bα1中称为Bar,Bβ1中称为Bbr)和3个信息素基因(Bαl中分别称为bapl(1)、bapl(2)和 bapl(3),Bβ1中分别称为bbpl(1)、bbpl(2)和bbpl(3)[4]。此外,玉米黑粉菌[5]和灰盖鬼伞的B交配型位点中都包含多个信息素基因及信息素受体基因。
1.2担子菌交配型基因的功能1.2.1A位点的交配型基因的功能A位点的大多数交配型基因都成对存在,并且由分别编码HD1和HD2的基因背向排列。在担子菌的性发育过程中,只有由来自不同位点的交配型基因编码的分别含HD1和HD2的蛋白质之间形成异源二聚体,才能完成调节作用,这一结论已得到许多实验结果的支持。实际上,两个基因之间发生亲和反应必须符合以下条件:(1)不同位点;(2)相同复合物;(3)相同基因对;(4)分别编码HD1和HD2[9]。A基因产物之间的二聚化是决定其功能的关键步骤。A位点的基因调控的细胞核配对、核的分裂和锁状细胞形成等发育过程与HD1~HD2异源二聚体有关。当两个可亲和的单核菌丝交配时,二者的HD1和HD2蛋白质相互识别并结合,具有转录因子的功能。显然,两种蛋白质间的识别非常重要,其交配特异性是由N-末端氨基酸序列决定的[10]。蛋白质的二聚化作用发生在细胞质中,作为转录因子,与DNA结合必须转移到核内,这依赖于HD1蛋白质中C-末端存在的核内定位信号。所以,HD2蛋白质的同源结构域提供DNA结合区域,HD1蛋白质则决定异源二聚体能否进入细胞核内。
1.2.2 B位点的交配型基因的功能真菌信息素受体基因编码的蛋白质二级结构很相似,都具有7个疏水的跨膜结构域(Transmembrane domain)[11],其作用是与信息素分子特异结合,并诱发下游基因的表达。真菌信息素基因编码的蛋白质中也存在保守结构,如C-末端都有CaaX或类似结构,其中C表示Cys,a表示脂肪族氨基酸,X表示任何一种氨基酸,CaaX是真菌信息素前体的特征结构,其作用是作为成熟过程中多肽C-末端进行异丙基修饰的信号,修饰后的信息素是高度疏水的。所有的真菌信息素基因都编码相当大的前体分子,然后被加工成9-15氨基酸的小肽,比较信息素的前体发现大都存在两个带电荷的残基,即ER/DR/EH/NH基序,这一部位可能是成熟过程中形成小肽的剪切点[12]。信息素与受体之间的反应是相当专一的,受体可以区分是否是由同一位点的基因编码的信息素,据信受体的专一性决定于其二级结构或三级结构。在担子菌中,对信息素反应途径的下游信息目前还知之甚少,但是与酿酒酵母(Saccha-romycescerevisiae)相比,在玉米黑粉菌中已发现各种中间产物的类似物,如Fuz7(Ste7p的类似物)[13]、Prf1(pheromoneresponsefactor,Ste12p的类似物)等,同时在灰盖鬼伞的信息素受体中发现了非常保守的序列[12]。这些都暗示担子菌中信息素—受体反应以及下游的应答与酵母中有类似之处。
1.3子囊菌交配型结构功能和命名子囊菌mt位点包含不连续的DNA序列,这些不连续的序列可能包含许多复合基因并且这些交配型基因具有明显的等位关系。子囊菌中有三个菌的个交配位点研究较深入,这三个菌(S.crassa、P.anserina和 C.heterostrostphus)交配位点包含相同的基因但是功能不同。N.crassa的交配型mt-a和mt-A最先研究清楚[6],mt-A长5.3kb包含三个基因分别称为mtA-1、mtA-2 和 mtA-3。mt-a长3.2kb只有一个转录单元mta-1。N.crassa的mt-a和mt-A中的侧翼不同。研究进一步表明,A位点的侧翼序列的存在不能影响a交配点的活性。 mta-1和mtA-1任何一个突变都将造成交配的障碍。但是只有mta-1能够转变交配型[7],是mta交配位点进行交配所必需的唯一基因。RIP(repeat–induced point mutation) 可被破坏mtA-1基因。对mta-1没有影响。mta-1的复制在有性生殖阶段发生,所以在双核融合时期mta-1必不可缺少。mtA-2和mtA-3在囊孢子的生成阶段必不可少。但是具体哪一个或者两个同时起作用还不清楚[1]。P.anserina中的mt等位点(idiomorph)与N.crassa的相似,mat+等位点只有一个转录单元FPR1,mat-等位点编码3个基因分别是FMR1、SMR2和SMR2。功能和N.crassa mtA的三个交配基因极为相似。这些交配型基因的的突变减少了交配几率,交配型基因的基本功能就是自我识别和非自我识别。当控制自我识别和非自我识别的基因缺失时,细胞核的形成主要通过无性繁殖产生。所以说,减数分裂和囊孢子的形成不能过分依赖于交配型基因。C.heterostrophus 的mt等位点每一个只含有一个交配基因。其中一个和S.cere-visiae 中的Matα1p是同源体,另一个和N.crassa中的mta-1是同源体。这两个交配基因对有性生殖的交配都起到同等重要的作用[8]。但是是否C.heterostrophus的mt位点外还具有和mta-2、mta-3相当的同源体至今还不清楚。以P.anserina的研究表明:并非所有的交配基因在mt位点必须存在,所以C.heterostrophus是否也具有和mt相似的交配位点也还不清楚。 3不发生交配时交配基因的功能有趣的是交配位点扮演的角色不仅仅是交配,包括生长亲和性、有性二态现象、子囊孢子二态现象和毒性等。这种现象是否和和编码的交配基因或与其相关的基因有联系目前还不能确定。交配基因的这些功能已经对种群的性状产生了深刻的影响。黑粉菌属更能体现这一点,因为其有性发育仅仅发生被感染的植株上。在菌丝生长阶段生长不亲和性是常见的,所以这样就限制了杂核体的形成。交配基因对担子菌杂核体亲和性的逆转起到重要作用,N. crassa交配位点有生长不亲和性的功能并阻止交配的发生。N. crassa至少有十个这样的位点,生长不亲和位点在功能上并不比交配位点更能阻止或加强性亲和。实际上,N. crassa的交配位点作为生长不亲和性功能不是很常见,甚至在真菌Neurospora中都很少见。当两个具有不同交配性基因的N. crassa(没有生长不亲和位点)在亲和情况下融合,菌丝死亡引起这种现象的分子机制至今还不清楚。这种现象可以人为创造,把含a的细胞质注射到含A的当中也会出现这种现象[14]。这种现象似乎说明配对交配基因在相遇之前在a细胞中存在一种因子使之发生不亲和反应。het-c、 het-e和 tol(tolenrant)位点都对生长不亲和反应起到不同程度的作用。tol位点的突变形成混合交配型异核体。tol位于染色体Ⅳ上,和trp-4紧密连接在一起[15],附加tol位点也可能存在,但是最新的研究发现仅有7个tol等位基因抑制子。交配基因可能还有其他的一些功能,但是这些方面的研究很少。
4交配型基因的进化除了关于交配分子机制的问题之外,交配的进化和群体遗传问题也是一个争论的热点。例如,交配系统是怎样出现的又是怎样表现出其多样性的?这些系统之间的变化带来什么样的优越性?交配系统和物种形成之间的关系如何?还有,交配系统和真菌无性生殖关系又怎样?这些问题的解决需要一段时间的研究才能慢慢解决。所有的交配系统之间都是相互关联的但并不是等同的,主要包括基因控制多肽和信号传导级联系统。由这些交配基因位点编码的“调节器装置”不是一承不变的[16],而是在不同的阶段相互变化的。例如:C. heterostrophus中含有MTa-1 和MTA-1同族体,并没有发现有MTA-2。S. cerevisiae和S. pombe也发现有更多的一基因多功能的现象。关于交配系统的起源和进化通过更多的研究物种之间的不同交配系统之间的关系可以更好的得以理解。更好的例子就是Neurospora,有异宗配合、假异宗配合和同宗配合物种。假异宗配合N. tetrasperma祖先来自于异宗配合物种Neurospora。因为最少又一个交配位点来自于N. crassa,实际上当外来基因渗入N. crassa中时, tol并不能阻止形成混合交配型异核体。同宗配合交配型不同,所有的物种都有一个基因序列在mtA位点杂交。但是只有N. terricola有一个mt位点的同族体。交配基因怎样在同宗配合物种中出现并不清楚,这些交配基因可能是异宗配合遗传物种留下的没有多大关系的剩余物。来自于同宗配合物种N. africana中的交配基因mtA-1与N. crassa有相同的序列,当被转入N. crassa时仍能起到mtA-1的特异性功能[17]。许多不同交配基因的不同组合可导致同宗配合现象(例如:两个交配基因的连锁或者编码区的融合)。关于交配基因的进化随着研究的深入将会逐渐的明了。目前研究的担子菌A位点和B位点都分别存在相似的结构特点。如灰盖鬼伞、裂褶菌和玉米黑粉菌都含有编码信息素及信息素受体的基因,越来越多的研究证实,信息素可能在真菌中普遍存在。与B位点相比,A位点的交配型基因的保守性更为广泛。灰盖鬼伞和裂褶菌的A位点[18]、玉米黑粉菌的b位点都是由一对或几对基因组成,每一对基因编码两种含相似同源结构域的蛋白质,在进化过程中,同源结构域序列是高度保守的,而且其氨基酸序列比相应的DNA序列保守程度更高,例如在灰盖鬼伞中,交配型基因b1-2和b1-1核苷酸的相似性为71%,而由此推出的相应多肽b1-2(642aa)和b1-1(681aa)氨基酸序列的相似性为79%[19]。这提示同源结构域是进化选择的结果。将担子菌中的这些交配型基因与子囊菌纲的酿酒酵母等进行比较,发现酿酒酵母的α2蛋白与HD1相似,a1蛋白与HD2相似[20]。事实上,目前已从动物、植物和菌类中克隆得到了大量的同源盒基因,其广泛存在说明了同源结构域在真核生物进化的早期就已出现。
5展望 担子菌和子囊菌的分子生物学研究近年来取得飞速,担子菌交配型基因的克隆大大加深了人们对传统真菌遗传学中A和B因子的认识,为剖析担子菌发育中交配控制的分子机理打下了基础。同源结构域的广泛存在及其保守性暗示其具有重要功能,同源结构域蛋白质特异区分不同来源的蛋白质形成异源二聚体,发挥调节作用,这不仅是真菌生物学家感兴趣的事情,而且对于高等真核生物的基因表达调控研究也具有重要意义。子囊菌亚门中,有很多重要的病原真菌,研究这些病原真菌的生活史及其有性生殖的交配基因的作用,将有利于更清楚的了解这些病原真菌,以便更好的控制这些病原真菌对农作物及作物造成的灾害。总之,真菌交配型基因的研究对真菌的改良及其控制(包括食用菌和病原菌)具有重要意义。 [1] J. W. Kronstad, C. Staben. mating type in filamentous fungi .Annu. Rev. Genet. 1997. 31: 245– 76[2] kues U,RichardsonWVJ,TymonAM etal.The combination of dissimilar alleles of the Aαand Aβgene complexs,whose proteins contain homeodomain motifs,determines sexual development in the mushroom Coprinus cinereus.Genes.Dev,1992,4:568—577.[3] Mating Type in Fungi (Molecular & Organismal Aspects) Plant Pathology 534 (Gen CB 534) Fungal Genetics Spring 2003[4]Vaillancourt IJ,RaudaskoskiM,SpetchCA etal.Multiple genes encoding pheromones and a phermone receptor define theBβ1 ,mating-type specificity in Schizophyllum commune.Genetics,1997,146:541—551.[5]BÖlkerM,Urban M and KahmannR.The a mating-type locus of Ustilago maydis specifies cell signaling components.cell,1992,68:441--450[6] 李莉云,刘国振,刘丽娟. 担子菌交配型基因的克隆及功能研究进展. 《生物工程进展》1999,Vol.19,No.1:19-23.[7]ChangS,StabenC.1994.Dirented replacement of mt a by mta-1 effects a mating type switch in Neurospora crassa.Genetics 138:75—81.[8] Turgeon BG, Bohlmann H, Ciuffetti LM,Christiansen SK, Yang G, et al. 1993.Cloning and analysis of the mating type genes from Cochliobolus heterostrophus. Mol. Gen. Genet. 238:270–84[9]李莉云,刘国振,贾建航.担子菌交配型位点结构及交配型基因功能的研究迸展,食用菌学报1998.5(2):57~63[10] Wu J,Ullrich RC and Novotny CP.Mol Gen Genet,1996,252:739-745.[11] VaillancourtLJ,Raudaskoski M,Spetch CA,etal.Genetics,1997,146:541-551.[12] O sheaSF,ChaurePT,HalshllJH.Genetics,inpress.[13] BanuettF,and HerskowitzI.GenesDev,1994,8:1367-1378.[14] Wilson JF, Garnjobst L, Tatuem EL.1961. Heterokaryon incompatibility in Neurospora crassa-microinjection studies.Am. J. Bot. 48:299–305[15] Newmeyer D. 1970. A suppressor of the heterokaryon-incompatibility associated with mating type in Neurospora crassa. Can. J. Gen. Cytol. 12:914–26[16] Nelson MA. 1996. Mating systems in ascomycetes: a romp in the sac. TrendsGenet. 12:69–74[17] Glass NL, Smith ML. 1994. Structure and function of a mating-type gene from the homothallic species Neurospora africana. Mol. Gen. Genet. 244:401–9[18] WuJ,UllrichRC and NovotnyCP.MolGen Genet,1996,252:739-745.[19] GieserPT and MayG. Gene,1994,146:167-176.[20] KuesU and CasseltonLA,Trends Genet,1992,8:154-156.
上一篇:关于现代中学数学教育的思考
下一篇:青枯菌病害的研究进展